蜜蜂群体是自然界中典型的集体行为之一,具有完善的功能特征和较强的适应性。本文通过梳理对蜂群行为及其决策过程的研究,将蜂群决策特点总结为:1.侦察蜂的组织方式提高了群体内知识的多样性;2.在参与决策过程时,侦察蜂没有表现出从众或盲目模仿他人的倾向;3.群体法定人数感知过程会以一种平衡决策准确性和速度这一对竞争需求的方式,聚集不同的和独立的意见。
来源: 集智俱乐部
作者:Thomas D. Seeley, Kevin Passino, Kirk Visscher
译者:汪显意
审校:梁金
编辑:邓一雪
几个世纪以来,社会选择问题一直困扰着社会哲学家和政治科学家们。群体基本的决策困境是,如何将个体对不同结果的偏好转变为群体作为整体的唯一选择。对这个问题的研究主要是关于人类群体,并发展出了各种各样的投票程序——多数人法则,相对多数人获胜,各种加权投票系统等——从一系列可能的选项中确认唯一的答案。对动物群体中的社会选择研究得较少,尽管例子十分丰富:一群狒狒必须决定休息后要去哪里,一个蚁群决定是否要攻击临近的蚁群。动物群体决策的一个代表性例子是,多达一万只蜜蜂组成的蜂群选择筑巢地点。这个过程涉及来自蜂群的数百只蜜蜂共同努力,在树丛中找到十几个或者更多的备选巢穴,然后从中选择一个最好的。在过去的十年里,我们一直在研究这个过程,使用各种观察、实验和数学建模方法。这项工作揭示了一系列群体的行为机制,群体可以据此长期作出出色的共同决策。很明显,这种群体智能是决策任务中代表不同选择的蜜蜂们之间分歧、争执的结果,而不是共识或妥协的产物。我们发现,演化,为群体如何作为一个有效的决策单元运转提供了一个有趣的答案。几个世纪以来,养蜂人知道,大型蜂群会在晚春或初夏分裂。在这个过程中,蜂后和约半数的工蜂会离开蜂巢,去建立一个新的蜂群。与此同时,新的幼年蜂后和余下的工蜂留下来维持旧的巢穴。养蜂人还知道,当蜂群离开亲代留下的蜂巢时,会在附近的树枝上聚成须状的簇,然后搜寻新的家园,即使是留在树枝上的蜜蜂,最终也会和蜂群一起离开,飞往它们的新居——通常是远处空心的树。长期以来,人们会捕捉临时驻扎在外面的蜜蜂,将它们安置在人造蜂巢中,缩短它们的巢穴搜索。因此,毫不奇怪,这个决策过程长期以来一直是一个深刻的谜团。20世纪50年代,当德国动物学家 Martin Lindauer 发表了关于蜜蜂寻房的开创性论文时,这种情况开始改变。Lindauer 当时是慕尼黑大学的博士后,与著名的 Karl von Frisch 一起学习,Karl von Frisch 不久前解码了蜜蜂的摇摆舞。这种沟通行为允许成功的觅食者通过一系列特定的动作告知同伴丰富的食物来源的位置。一只跳舞的蜜蜂向前跑并且摇摆,横向振动腹部,然后绕回起点,产生一个舞蹈回路。一次舞蹈包含许多这样的回路。Von Frisch 发现,蜜蜂摇摆跑动的长度可以翻译成到食物来源的距离,舞蹈的角度代表食物的方向。Lindauer 是一个敏锐的观察者。有一次,当他将自己的技能用于在大学动物研究所外定居的一群蜜蜂时,他注意到蜂群表面的蜜蜂正在表演摇摆舞。此外,他观察到,与蜂巢里的蜜蜂不同,这些舞者没有携带大量的花蜜或花粉。显然,这些不是宣传食品来源的觅食者。他们可能是报告潜在巢穴地点的侦察兵吗?这是以前未知的摇摆舞用法。图3. 蜜蜂跳摇摆舞来告知食物来源,也可以告知巢穴的位置。蜜蜂离巢时,花沿着太阳右侧40度的一条线排列(左上)。为了报告这个食物来源,一只蜜蜂在蜂巢上跑出数字8的图案(右上)。当她穿过舞蹈的中心部分时,表演摇摆奔跑,横向振动她的身体,奔跑的角度表明了食物来源的方向。摆动的持续时间与食物来源的距离有关(底部)。当摇摆舞指示的是巢穴时,它发生在蜂群表面,而不是在蜂巢内。| 图片:Stephanie FreeseLindauer 耐心地观察了几个蜂群中的所有舞者,回答了这个问题,这是项艰巨的任务,需要许多天稳定地观察蜜蜂和疯狂地做笔记。每当他看到一只新的跳舞的蜜蜂时,Lindauer 都会关注她的舞蹈所编码的位置,然后给她画一个点标记,以避免反复记录她的舞蹈信息。这项艰苦的工作收获了几个了不起的发现。一个是,在决策过程中,蜂群里数千只蜜蜂中只有几百只是活跃的——在蜂群里飞进飞出,大概是寻找潜在的筑巢地点,然后舞蹈。大多数蜜蜂保持静止,可能是为了保护蜂群的能量供应,直到蜂群做出决定,再飞往选定的地点。第二个奇怪的发现是,起初,蜜蜂的舞蹈指示周围的许多地点,但随着时间一小时一小时过去,宣传的地点数量逐渐下降,直到只剩下一个,数十只跳舞的蜜蜂兴奋地公布这个地点。Lindauer还发现,在蜜蜂的舞蹈指向一个地点后不久,整群蜜蜂就会突然起飞并飞向这个地点。有时,在蜂群的整个越野飞行中,他跟在下面狂奔,从而了解它们确切的目的地——通常是树或建筑物上的空洞,并且总是在最后舞蹈中指明的地方。毫无疑问,跳舞的蜜蜂是在报告筑巢地点。事实上,这些蜜蜂似乎正在就蜂群的未来家园举行某种公民投票,尽管他们到底是如何进行商量的仍然不得而知。20世纪90年代中期,我们决定更深入地研究这个有趣的动物民主的例子。在 Lindauer 的工作之后的几年里,几位研究者考察了蜜蜂的住址偏好,发现对于蜂群而言,头等家园有体积大于20升的空腔;入口孔小于30平方厘米,离地面几米,朝南,位于空腔底部。但没有人确切知道蜂群中的侦查蜂(scout bees)在集体选择新家时是如何实施这些住房偏好的。我们重新分析的第一步是重复 Lindauer 对侦查蜂舞蹈的观察,但使用现代录像设备来获得比20世纪50年代更完整的画面。我们观察一个大约有4000只蜜蜂的小蜂群,给每只蜜蜂贴上标签以便识别,这样我们就可以将每支舞蹈归因于特定的个体,从而确定她对蜂群决策的贡献。图4. 标记的蜜蜂。| 照片:Thomas D. Seeley从每只侦查蜂的每支舞蹈的录像中,我们发现了一种巢穴侦察兵的舞蹈模式,这与 Lindauer 根据他仅对每个侦查蜂的第一次舞蹈的记录所做的报告非常相似。例如,1997年7月20日至22日期间,在我们观察的蜂群中,整个决策过程需要在3天里进行约16个小时的舞蹈活动。在这个过程的前半部分,侦察兵们报告了他们将考虑的所有11个潜在巢穴地点,没有一个地点主导舞蹈。然而,在下半场,其中一个地点逐渐开始有比其他地点更多的宣传,并最终成为被选中的地点。事实上,在决策的最后几个小时里,作为领跑者的地点成为所有舞蹈的对象。
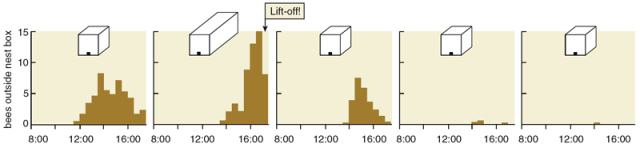
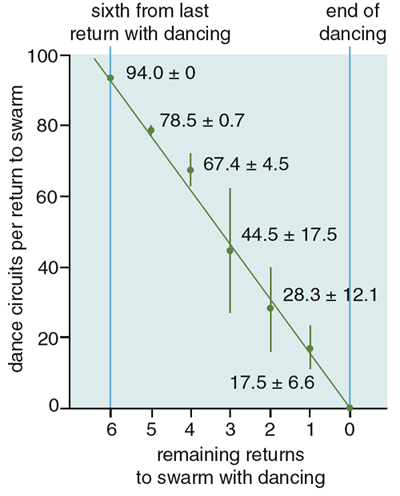
鉴于蜂群中的舞蹈代表一个地点,然后蜂群移动到这个地点这一惊人方式,人们很容易地得出结论:蜂群的决策过程本质上是建立共识的过程,就像贵格会教徒之间“会议的感觉”(Sense of the Meeting)的到来一样。根据这个假设,一只侦察蜂通过跳舞来为一个地点“投票”,这些侦察蜂以某种方式行动和互动,以便他们的投票逐渐达成一致,支持一个优越的地点,并且由于某种原因投票模式受到稳步监控,以便每只侦察蜂知道它们何时达成一致,并根据自己的决定采取行动。图5. 蜂群的决策过程示意图。对不同地点的投票逐渐达成一致,支持最优越的地点。| 图片:Thomas D. Seeley然而,有两个因素使人们对这个有吸引力的假说产生了怀疑。首先,Lindauer 和我们都没有观察到任何侦察蜂给它们的舞者同伴投票的迹象,但这是它们肯定要做的,用来知道什么时候达成共识。其次,Lindauer 和我们都会偶尔看到蜂群在没有舞蹈共识的情况下起飞,也就是说,此时有两个强大的舞者联盟为两个不同的地点宣传。这些罕见的持不同意见起飞的案例是我们可以忽视的异常,还是我们应该注意的宝贵线索?我们选择留意他们,因为我们长期以来一直想知道,蜂群决策的本质或许是在其中一个地点感知到法定人数(sensing a quorum)(足够数量的侦查蜂),而不是在蜂群集群中感知共识(sensing a consensus)(跳舞的侦察蜂的共识)。根据这个法定人数感知假说(quorum-sensing hypothesis),侦察蜂通过待在这个地点为它“投票”,侦察蜂以某种方式行动和互动,使他们在优越地点的数量增长得更快,每个地点的蜜蜂都会以某种方式监控这个地点的蜜蜂总数,以便知道它们是否达到阈值数字(法定人数),并开始发动蜂群迁移到这个地点。这个假说可以解释蜂群意见不统一就起飞的情况,即在不同地方的舞者之间的竞争只留下唯一一个地点而排除其他选项之前,一个地点的蜜蜂就达到了阈值数量的情况。我们在康奈尔大学浅滩海洋实验室(Shoals Marine Laboratory of Cornell University)所在地阿普尔多尔岛(Appledore Island)进行了实验来测试这两个假说。这个岛远离缅因州海岸,上面几乎没有树木,对于蜜蜂来说缺乏天然的适合筑巢的空腔。因此,我们运送到这个岛上的每个蜂群都被迫注意我们提供的特殊巢箱。图6. 实验场景。| 图片:Thomas D. Seeley在第一个实验中,我们测试了几个蜂群,一次一个,有两个相同的巢箱,每一个都是绝佳的筑巢地点。蜂群被放置在岛的中央,两个巢箱都放置在岩质海岸附近,每个巢箱距离蜂群250米,但方向不同。我们发现,当蜂群被迫在两个一流的巢穴中进行选择时,它们通常会在侦察蜂还为两个地点强烈地跳舞时就起飞。对于蜂群开始飞往其中一个地点,舞者之间的共识无疑是非必要的,因此我们可以否定共识感知假说(consensus-sensing hypothesis)。与此同时,我们得到了对法定人数感知假说的支持,因为我们注意到,一旦在一个巢箱里看到15只或更多蜜蜂在一起,蜂群就开始为飞行做准备。然而,应该指出的是,由于蜜蜂大部分时间都在蜂群中度过,任何时候在巢穴中看到至少15只蜜蜂就意味着总共大约有150只蜜蜂正在访问该地点。在阿普尔多尔岛的第二次实验中,我们通过验证一个可以被检验的预测,明确测试了法定人数感知假说:延迟蜂群选择的筑巢地点处法定人数的形成,同时让其余决策过程不受干扰,将会推迟群体到该地点的飞行。为了推迟法定人数的形成,我们在岛上的一个地方将五个理想的巢箱放在一起。这导致访问该地点的侦察蜂分散在五个相同的巢穴中,而不是集中在一个巢穴。然后,我们看到,从发现巢箱的位置开始,蜂群花了多长时间才做出决定并起飞飞往那个地点。我们还对每个蜂群进行了一次仅有一个巢箱的控制试验。每个蜂群的两次实验都是使用岛上的两个不同地点进行的,因此每次试验都以同样的方式开始:一只侦察蜂在一个新地点发现了一个有吸引力的巢穴。在我们测试的所有四个群体中,与单巢箱组(平均196分钟)相比,五巢箱组中确实存在显著的起飞延迟(平均442分钟)。因此,该实验为法定人数感知假说提供了强有力的支持。图7. 第二个实验的结果。| 图片:Stephanie Freese侦察蜂具体如何感知法定人数仍然是一个谜。他们可能会使用视觉、嗅觉甚至触觉信息来估计地点的侦察蜂数量,但这仍然是未来研究的主题。一旦一个地点中的蜜蜂数量达到法定人数阈值,蜜蜂就会开始一种众所周知的行为。该地点的侦察蜂将返回蜂群集群,并开始产生特殊的高频率的声学信号,刺激蜂群集群中的非侦察蜂们开始通过颤抖将飞行肌肉加热到飞行所需的33至35摄氏度。在产生这个信号(我们称之为工人管道(worker piping))时,一只侦察蜂爬行穿过蜂群,每隔一秒钟左右停下来,将她的胸部压在另一只蜜蜂身上,并激活她的翅膀肌肉。虽然大多数振动能可能直接转移到她接触的蜜蜂体内,但这种动作会产生一种可听到的振动,让人想起赛车发动机的转动。管道信号持续约0.8秒,基本频率约为200赫兹。由于对工人管道的刺激是所选地点的侦察蜂的法定人数,而不是所有侦察蜂对该地点的共识,因此群体变暖过程通常在侦察蜂达成共识之前就开始了。但由于热身通常需要一个小时或更长时间,所以在整个蜂群起飞之前,侦察蜂通常有时间就所选地点达成共识。图8. 侦察蜂感知法定人数的过程。| 图片:Thomas D. Seeley, Stephanie Freese通过观察侦察蜂的舞蹈来窃听蜂群的决策,Lindauer 和我们的团队清楚地表明,蜂群从一批五个或更多选项中选择一个筑巢地点。自然产生的下一个问题是,蜂群选择的是否是最佳地点,如果是,如何做到的?为了评估蜂群选择巢穴的准确性,我们向阿普尔多尔岛的蜂群提供了五种选择,其中四种选择是质量中等的巢箱,一种是质量一流的巢箱。四个一般的巢箱在各方面都很有吸引力,除了每个箱子只提供15升的生活空间外。最好的巢箱与其他四个完全相同,只是它提供了40升的空间,这个体积更好地满足了各种聚居活动(繁殖、储存食物等)对空间的需求。图9. 蜜蜂选择最佳蜂巢的过程。| 图片:Thomas D. Seeley几乎所有的测试蜂群都选择了优质的巢箱。具体而言,我们观察到,尽管优质巢箱从不是第一个被发现的地点,但一旦侦察蜂发现了这个最理想的地点,来到该地点的蜜蜂数量就开始增长得比其他所有地点都快,并首先达到法定人数阈值。此外,随着优质地点蜜蜂数量的增加,每个一般地点的蜜蜂数量都有所减少,这表明对最佳地点的兴趣的不断上升会降低对其他地点的兴趣。这种通过在最佳地点增强来抑制在较差地点聚集的机制十分重要,因为它有助于确保在最佳地点首先超过法定人数阈值,并在侦察蜂之间产生共识模式,这种模式几乎总是在蜂群飞向新家前不久出现。这种动力学背后的单个侦察蜂层面的行为机制是什么?其中之一是,侦察蜂对舞蹈力量的精细调整,即它们为一个地点表演的摇摆舞回路的数量,是该地点质量的函数。我们研究了这种现象。我们在阿普尔多尔岛给一个蜂群同时呈现两个巢箱,一个优质的,一个一般的,并分析蜂群在两个巢箱的摇摆舞。我们发现,当第一次有侦察蜂从最好的的地点回到蜂群时,她倾向于表演包含100个或更多舞蹈回路的摇摆舞。侦察蜂们也报告一般但可以接受的筑巢地点,大概是以防没有更好的地方。但当一个侦察蜂第一次从某个一般的地点回来时,我们发现她可能会表演一个只有十几个舞蹈回路的摇摆舞。为特定地点跳舞的力量越大,新来者的流量就越大,因此在最佳地点,侦察蜂的积累速度会最快。来自很好和一般地点的侦察蜂的招募信号强度之间的差异,被它们行为的另一个奇怪特征放大。如果一只侦察蜂飞往一个地点,她将多次飞往该地点(可能既是为了支持“她的地点”,也是为了随时了解那里的侦察蜂同伴数量),每次访问后,她都会用摇摆舞为她的地点做宣传。不过,每次她回到蜂群表演舞蹈时,都会将舞蹈广告的强度减少约15个舞蹈回路。其结果是,两个站点之间招募信号强度的总体差异几乎是站点之间质量差异的指数函数。如果宣传优质地点和一般地点的两只蜜蜂在第一次回到蜂群时分别表演90个和30个舞蹈回路,那么它们的招募信号的总差异不仅仅是三倍,而是七倍(90 + 75 + 60 + 45 + 30 + 15 + 0 = 315个回路,对 30 + 15 + 0 = 45 个回路)。此外,在宣传过程中存在很强的正反馈,因此到一个地点的蜜蜂数量越多,招募到的蜜蜂数量就越多,这反过来又导致来到该地点的蜜蜂数量更多。因此,两个筑巢地点之间质量和摇摆舞强度的微小差异,可能会使与这些地点相关的侦察蜂数量相差巨大。图10. 侦察蜂每次回到蜂群时,会将舞蹈强度减少约15个舞蹈回路。| 图片:Stephanie Freese摇摆舞力量的差异和招募过程固有的正反馈,解释了致力于不同候选地点的侦察蜂数量的差异,最好的地点获得侦察蜂的速度最快。但是,是什么导致较差地点的支持者数量急剧下降,而较好地点的支持者数量却急剧上升呢?为劣质地点宣传的侦察蜂数量下降的根本原因是,所有侦察蜂,即使是那些致力于优质地点的侦察蜂,最终都会放弃他们的站点。通常,蜜蜂在停止为一个地方舞蹈后不久就停止访问该地点,因此蜜蜂离开差的地方比离开好地方更快。一旦一只侦察蜂放弃一个地点,她就会“重置”,并可以被招募到另一个地点,甚至被重新招募到同一个地点。然而,当蜜蜂为一个地点跳完舞时,大约80%的时间她会完全停止跳舞。因此,侦察蜂依赖于招募其他侦察蜂,这些侦察蜂在搜索中找不到任何候选地点,因此不致力于宣传任何地点。但是,当蜜蜂被招募访问一个地点时,如果她觉得这个地点很差,她可能不会在回来时立即为该地点跳舞。因此,一个未跳舞的侦察蜂可能会在找到她认为有价值的地点之前访问好几个地点。图11. 侦察蜂在选择巢穴时的数量变化过程。| 图片:Stephanie Freese只要一个地点的招募率超过放弃率,隶属于该地点的侦察蜂数量就会增加。不论如何,最终,最优质地点的招募率将滚雪球般增大,到那时,每个劣势地点的招募率将逐渐消失:没有为某个地点跳过舞的侦察蜂数量是有限的,大多数都被招募到最佳地点。当每个劣势地点的招募率低于放弃率时,致力于这些地点的侦察蜂数量开始减少。简而言之,随着致力于最佳地点的群体变大,它会自动将隶属于劣等地点的团体排除在竞争之外。悉尼大学(University of Sydney)的数学生物学家 Mary R. Myerscough 创建了在不同巢穴表演舞蹈的侦察蜂数量动力学的数学模型。她优美地证明,只要有足够的时间,成群结队跳舞的侦察蜂几乎总是会专注于已经找到的最好的地点。这当然符合 Lindauer 和我们在侦察蜂的辩论中看到的情况:在蜂群起飞飞回家之前,舞者之间几乎总是会达成共识。尽管在蜂群的舞蹈记录中,在起飞前舞蹈者之间的共识是一个显著的特征,但我们现在明白,达到法定人数,而不是建立共识,是蜜蜂群体决策过程的本质。然而,我们不应该将舞者共识视为蜜蜂决策过程的不重要的副产品。相反,为了让蜂群成功飞往新家,达成共识是必要的。我们偶尔会看到,当一个蜂群起飞时,侦察蜂还在为多个地点跳舞,每次,空中的蜂群都无法飞走。蜂群飞行引导机制尚不清楚,但从这些观察中可以清楚地看出,引导过程依赖于足够数量的侦察蜂向其他飞行的蜂群提供一致的方向信息。当分裂决定发生时,蜂群似乎需要重新停下并继续审议,直到一个地点占主导地位。任何决策者面临的一个基本问题是,在快速决策和良好决策之间找到合适的折中方案。如果一个动物或一个群体必须迅速做出决定,那么它很容易做出糟糕的决定,因为它要么不能对自己的选择进行足够广泛的采样,要么无法对它们进行足够深入的评估,或者这两点都有。假设蜜蜂群在选择筑巢地点时在速度和准确性之间经过了这样的权衡,我们想知道,蜜蜂群体决策过程的行为参数是否经过自然选择的调整,使得蜂群在减少时间和能量成本的同时,最大限度地减少选择不良地点的概率。为了验证事实是否如此,我们建立了一个随机的、离散的蜂群决策过程的数学模型,然后使用我们的模型来创建“伪突变”蜂群,这些蜂群对各种行为参数具有不同的值。这使我们能够看到特定参数的增加或减少如何影响蜂群选择巢穴的速度和准确性。一个明显的候选参数是法定人数大小,因为法定人数感知是蜂群决策的核心。当我们改变模型中的法定人数,同时将其他一切参数都保持在正常水平时,该模型清楚地表明,低法定人数会产生相对快速但通常不准确的决定,而高法定人数会产生更慢但更准确的决策。特别值得注意的是,该模型预测的,能很好地平衡速度和准确性的法定人数规模,大约是15至20只蜜蜂,基本上与经验性的发现相吻合,即当一个地点的蜜蜂数量达到10到20只时,侦察蜜蜂就会启动蜂群变暖过程,为起飞做准备。我们还研究了侦察蜂行为的一个奇怪特征,该特征可能有助于蜂群的决策,即侦察蜂在反复访问一个潜在的巢穴地点时,会降低她跳舞的强度。令人惊讶的是,每当一只侦察蜂访问一个潜在的巢穴地点,然后返回蜂群为这个地点做广告时,她跳的摇摆舞就会比之前有更少的舞蹈回路,因此对她的地点的宣传越来越不强烈。我们模型中舞蹈回路减少率的改变揭示了这个因素对决策过程有多重要。在模型中,改变舞蹈回路减少的速度,揭示了这个因素对决策过程的重要性。如果舞蹈回路数量减少的速度比在自然界中观察到的要快,那么做出决定所需的时间就会稳步增加,因为回路数量的快速衰减使得蜂群很难在任何一个地点达到法定人数。相反,如果回路数量的减少速度比自然界中观察到的要慢,那么就会出现一个更大的问题:随着分裂决策(即在多个地点快速达到法定人数)变得普遍,蜂群决策会完全失败。同样值得注意的是,我们的模型对舞蹈回路减少速率的预测在速度和准确性之间提供了一个很好的平衡——每次访问一个筑巢地点15到20个舞蹈回路——基本上符合经验发现,平均而言,侦察蜂每次访问一个巢点会缩短15个舞蹈回路。鉴于这些关于法定人数大小、舞蹈回路减少速度和其他参数的发现,我们得出结论,蜜蜂群中侦察蜂的行为确实经过了自然选择的调整,以创建一个在速度和准确性的竞争需求之间取得有利平衡的群体决策过程。
亨利·大卫·梭罗(Henry David Thoreau)在他1838年的一篇日记中哀叹,人类群体在实现集体智慧方面遇到了困难:“大众永远不会达到它最好的成员的标准,相反,它会贬低自己到最低的水平。”同样,弗里德里希·尼采(Friedrich Nietzsche)在《超越善恶》中写道:“愚蠢的行为对个人来说是例外,但对群体来说却是常态。”虽然群体确实会做出不好的决定,但群体也能做出好的决定。在什么样的情况下,群体才能具有高度的智慧,并能够合作做出正确的选择?我们认为,蜜蜂的巢址选择行为可以为这一主题提供指导,因为很明显,它们成功地做出了集体判断。第一个相关的因素是,侦察蜂的组织方式提高了群体内知识的多样性。具体来说,它们不是由少数蜜蜂领导或主导的;相反,决策过程在蜂群中广泛传播。因此,蜂群的决策过程是基于数百个个体的行动,每个个体都是一个自治的主体,能为解决找房子问题提供独特的信息。例如,请注意蜜蜂如何完成决策任务的第一阶段——发现可能的选择。一个蜂群中的几百只侦察蜂独立、广泛、同时地搜索,为蜂群带回各种各样的信息——优质的、一般的、甚至糟糕的地点——这些信息可以通过摇摆舞的方式与其他侦察蜂分享。所有潜在巢穴的发现都是自由报道的;没有侦察兵被压制。因此,蜂群充分利用其固有的群体特性,迅速地聚集起——通常只需要几个小时——大量选择。这个选择的集合越大,就越有可能包含一流的地点。因此,我们看到,蜂群决策的一个关键特征是它的分散组织,这有助于确保它有广泛的选择集合。蜜蜂行为中促进群体智能的第二个特征是,在参与决策过程时,侦察蜂没有表现出从众或盲目模仿他人的倾向。我们已经解释过,这个过程的核心是隶属于不同地点的各种侦察蜂联盟之间的竞争,每个联盟都在竞争吸引没有跳舞的侦察蜂到她的站点。每个联盟的成员通过表演摇摆舞来招募额外的成员,摇摆舞的强度与地点质量有关,这样地点质量越高,摇摆舞就越强,新来者的流量就越大。这里至关重要的是,当一个未跳舞的侦察蜂被招募到一个地点时,她不会盲目支持那只带她来的蜜蜂。相反,她会自己考查宣传的地点,只有当她也判断这是一个有价值的地点时,她才会为它跳舞,从而吸引更多的蜜蜂到这个地点。通过这种观点的独立性,侦察蜂避免了在评估地点时传播错误。只有在真正好的地点,舞者才能吸引更多的舞者,因此,在该地点的侦察蜂的数量将会有很大的增加。最终结果是,侦察蜂避免了对不良选择的群体狂热。群体成功的第三个关键是,群体法定人数感知过程会以一种平衡决策准确性和速度这一对竞争需求的方式,聚集不同的和独立的意见。法定人数水平足够高,许多蜜蜂在选择地点之前必须独立评估其质量。仅根据一只或几只蜜蜂的良好评价来快速选择一个家是不可能的。法定人数感知过程过滤掉极端的或不准确的意见,并对所选地点提供平衡的、群体级别的评估。这个评估过程需要时间,但确保有足够的时间间隔来产生真正的意见多样性,并确保在选定其中一个地点之前对所有已发现的地点进行了独立地评估。因此,汇总蜜蜂信息的法定人数感知方法允许意见的多样性和独立性很好地发展,但时间也足够长,来保证决策错误不太可能发生。这些考虑说明了蜜蜂群体决策的研究如何帮助人类群体实现集体智慧,从而避免集体愚蠢。蜜蜂向我们表明,良好的群体决策可以通过赋予一个群体三个关键的习惯来培养:将每次商议构建为公开的思想竞争;促进群体成员之间的知识多样性和意见独立性;并以一种符合时间限制的方式汇总意见,同时明智地利用群体内部知识的广度。https://www.americanscientist.org/article/group-decision-making-in-honey-bee-swarms
Beekman, M., R. L. Fathke and T. D. Seeley. 2006. How does an informed minority of scouts guide a honey bee swarm as it flies to its new home? Animal Behaviour 71:161-171.Black, D. 1986. The Theory of Committees and Elections. Dordrecht: Kluwer.Camazine, S., P. K. Visscher, J. Finley and R. S. Vetter. 1999. House-hunting by honey bee swarms: collective decisions and individual behaviors. Insectes Sociaux 46:348-360.Conradt, L., and T. J. Roper. 2005. Consensus decision making in animals. Trends in Ecology and Evolution 20:449-456.Franks, N. R., S. C. Pratt, E. B. Mallon, N. F. Britton and D. J. T. Sumpter. 2002. Information flow, opinion polling and collective intelligence in house-hunting social insects. Philosophical Transactions of the Royal Society of London B 337:1567-1583.Lindauer, M. 1955. Schwarmbienen auf Wohnungssuche. Zeitschrift für vergleichende Physiologie 37:263-324.Myerscough, M. R. 2003. Dancing for a decision: A matrix model for nest-site choice by honey bees. Proceedings of the Royal Society of London B 270:577-582.Passino, K. M., and T. D. Seeley. 2006. Modeling and analysis of nest-site selection by honey bee swarms: The speed and accuracy trade-off. Behavioral Ecology and Sociobiology 59:427-442.Seeley, T. D. 2003. Consensus building during nest-site selection in honey bee swarms: The expiration of dissent. Behavioral Ecology and Sociobiology 53:417-424.Seeley, T. D., and S. C. Buhrman. 1999. Group decision making in swarms of honey bees. Behavioral Ecology and Sociobiology 45:19-31.Seeley, T. D., and S. C. Buhrman. 2001. Nest-site selection in honey bees: How well do swarms implement the "best-of-N" decision rule? Behavioral Ecology and Sociobiology 49:416-427.Seeley, T. D., and J. Tautz. 2001. Worker piping in honey bee swarms and its role in preparing for liftoff. Journal of Comparative Physiology A 187:667-676.Seeley, T. D., and P. K. Visscher. 2003. Choosing a home: How the scouts in a honey bee swarm perceive the completion of their group decision making. Behavioral Ecology and Sociobiology 54:511-520.Seeley, T. D., and P. K. Visscher. 2004. Quorum sensing during nest-site selection by honey bee swarms. Behavioral Ecology and Sociobiology 56:594-601.Surowiecki, J. 2004. The Wisdom of Crowds. New York: Doubleday.
未来智能实验室的主要工作包括:建立AI智能系统智商评测体系,开展世界人工智能智商评测;开展互联网(城市)大脑研究计划,构建互联网(城市)大脑技术和企业图谱,为提升企业,行业与城市的智能水平服务。每日推荐范围未来科技发展趋势的学习型文章。目前线上平台已收藏上千篇精华前沿科技文章和报告。
如果您对实验室的研究感兴趣,欢迎加入未来智能实验室线上平台。扫描以下二维码或点击本文左下角“阅读原文”

关注公众号:拾黑(shiheibook)了解更多
[广告]赞助链接:
四季很好,只要有你,文娱排行榜:https://www.yaopaiming.com/
让资讯触达的更精准有趣:https://www.0xu.cn/
















 人工智能学家
人工智能学家
 关注网络尖刀微信公众号
关注网络尖刀微信公众号





